
We are continuing our efforts to recapitulate oxidative protein folding in vitro: in particular by developing efficient systems for folding proteins with complex disulfide connectivities. The insights gained should help inform current debate concerning the way disulfide bonds are generated in the endoplasmic reticulum of higher eukaryotes.
We have found that riboflavin binding protein (RfBP) is a useful oxidative protein folding substrate. RFBP has 9 disulfides (and consequently 33 million disulfide isomers for the fully oxidized protein). A key advantage is that the native apoprotein binds riboflavin tightly and rapidly with complete quenching of the flavin fluorescence. This allows us to follow oxidative protein folding continuously.
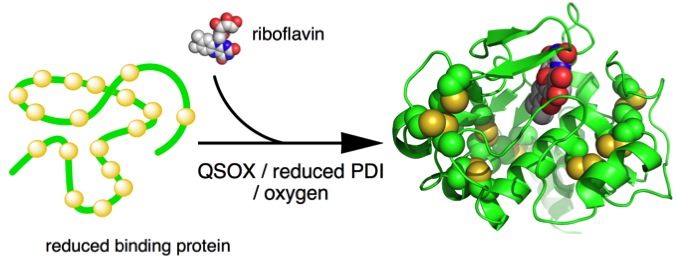 We have shown that efficient oxidative refolding of RfBP can be achieved with nanomolar levels of QSOX, and concentrations of reduced PDI that are realistic for the endoplasmic reticulum. Neither oxidized PDI nor glutathione are necessary for rapid folding.
We have shown that efficient oxidative refolding of RfBP can be achieved with nanomolar levels of QSOX, and concentrations of reduced PDI that are realistic for the endoplasmic reticulum. Neither oxidized PDI nor glutathione are necessary for rapid folding.
In this model system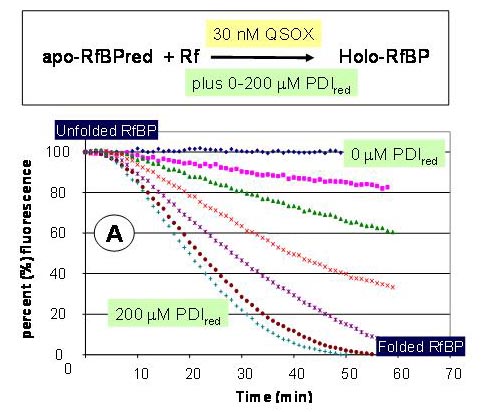 we can also dispense with QSOX and use mixtures of reduced and oxidized PDI as a redox buffer without other redox components (in particular oxidized or reduced glutathione). The fastest regain of riboflavin binding ability comes when the ratio of reduced/oxidized PDI is the largest (a little oxidized PDI is necessary for the stoichiometric oxidization of the 9 disulfides in native RfBP). It seems likely that the optimal ratio of reduced/oxidized PDI may be dependent on the nature, and particularly disulfide complexity, of the client protein. Nevertheless, in this case, refolding occurs most efficiently at the most reducing of PDI redox poises.
we can also dispense with QSOX and use mixtures of reduced and oxidized PDI as a redox buffer without other redox components (in particular oxidized or reduced glutathione). The fastest regain of riboflavin binding ability comes when the ratio of reduced/oxidized PDI is the largest (a little oxidized PDI is necessary for the stoichiometric oxidization of the 9 disulfides in native RfBP). It seems likely that the optimal ratio of reduced/oxidized PDI may be dependent on the nature, and particularly disulfide complexity, of the client protein. Nevertheless, in this case, refolding occurs most efficiently at the most reducing of PDI redox poises.
Since both reduced PDI is an extremely poor substrate of QSOX – and QSOX works best with unfolded reduced proteins – then QSOX can selectively insert disulfides into client proteins leaving reduced PDI to isomerize the mistakes it makes. If this were to operate in the endoplasmic reticulum, collateral oxidation of the glutathione redox pool would also be avoided – since the primary oxidation of cliient proteins occurs via QSOX, and not oxidized PDI.
The extent to which QSOX and reduced PDI cooperate in vivo is currently unknown.
