
Depending on the source, QSOX sequences start with one or two thioredoxin domains (the bar diagram is for metazoans). The remaining two domains are common to all QSOXs: a helix-rich region (HRR) and a flavin-binding domain (Erv/ALR, yellow). The latter is the engine of QSOX catalysis – driving the generation of disulfide bonds and reducing oxygen to hydrogen peroxide. In addition to QSOX, this Erv/ALR domain is found in a number of small dimeric stand-alone sulfhydryl oxidases – exemplified by the structure of yeast Erv2p [PubMed].
The origins of QSOX lie deep in the eukaryotic tree – before the divergence of the fungi. So where did that mysterious HRR domain come from? Our sequence analyses were uninformative.
In a recent paper, the Fass laboratory (Alon et al.) show that the HRR domain from human QSOX1 bears a remarkable structural resemblance to the flavin binding domain of Erv/ALR itself. Even the relative alignment of helices between the HRR and Erv/ALR domains have been preserved from the dimeric Erv2p protein.
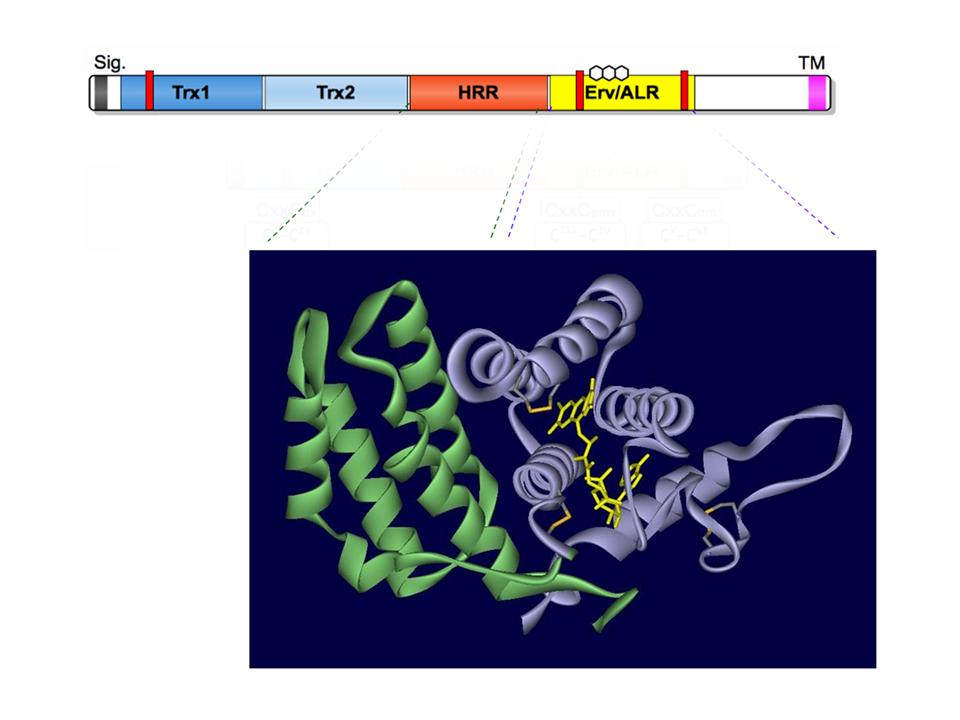 Gene duplication of the 4-helix-flavin-binding-bundle (a signature of these sulfhydryl oxidase structures) was seemingly followed by a loss of FAD binding determinants and the emergence of the flavin-free HRR domain. The combination of thioredoxin, and active and degenerate Erv/ALR domains led to the proficient QSOX catalysts that we study today.
Gene duplication of the 4-helix-flavin-binding-bundle (a signature of these sulfhydryl oxidase structures) was seemingly followed by a loss of FAD binding determinants and the emergence of the flavin-free HRR domain. The combination of thioredoxin, and active and degenerate Erv/ALR domains led to the proficient QSOX catalysts that we study today.
Now, where did the Erv/ALR flavin-binding domain (shown in purple above) come from?
