
What’s your favorite element? – for many scientists studying oxidative protein folding it would have to be sulfur. Not only does the oxidation of cysteinyl sulfur generate the constellation of structural disulfide bonds found in secreted proteins, but cysteine residues are an essential catalytic ingredient in the enzymes that form and isomerize these very disulfide linkages. While carbon, nitrogen and hydrogen can be routinely probed by nuclear magnetic resonance, the application of biological NMR to sulfur is impractical – there is no suitable sensitive NMR-active isotope for this critical player in oxidative protein folding.
Now Sharon Rozovsky and her colleagues have devised a simple way to substitute the NMR-active 77Se isotope for sulfur in recombinant proteins [PubMed]. Escherichia coli cells are grown in minimal media until they approach sulfur starvation, and then the medium is supplemented with ratios of Se (as selenite) to S (as sulfate) that are needed to achieve the desired incorporation ratio of Se/S. Anyone who expresses recombinant proteins in minimal media should be able to make this method work following the protocol outlined in Schaefer et al. [PubMed].
One of the proteins they have studied is augmenter of liver regeneration (ALR) – an enzyme that drives disulfide bond generation in the intermembrane space of the mitochondrion. The short form of ALR is a covalent homodimer (32 kDa) containing four structural disulfide bonds, two catalytic disulfides, and two FAD prosthetic groups. Using the random selenium incorporation method of Rozovsky and coworkers, Se/S ratios of up to 90% are readily achievable for human ALR. The resulting Se-ALR enzyme is almost as stable as the wild type protein, and the substituted protein retains significant enzymatic activity. Importantly, multiple selenocysteine and selenomethionine resonances can now be visualized by conventional solution NMR without the need for high field strengths or lengthy acquisition times.
But what are the structural consequences for such a wholesale substitution of sulfur for selenium in ALR? Schaefer et al. were able to crystallize the 90% substituted protein and compare it to the coordinates of the human ALR protein already solved in the laboratory of Brian Bahnson [PubMed]. On the left you can see the structure of Se-ALR refined to a resolution of 1.5 Angstroms with an occupancy of 90% Se / 10% S [PDB link] . The protein fold is essentially identical to that of native human ALR.
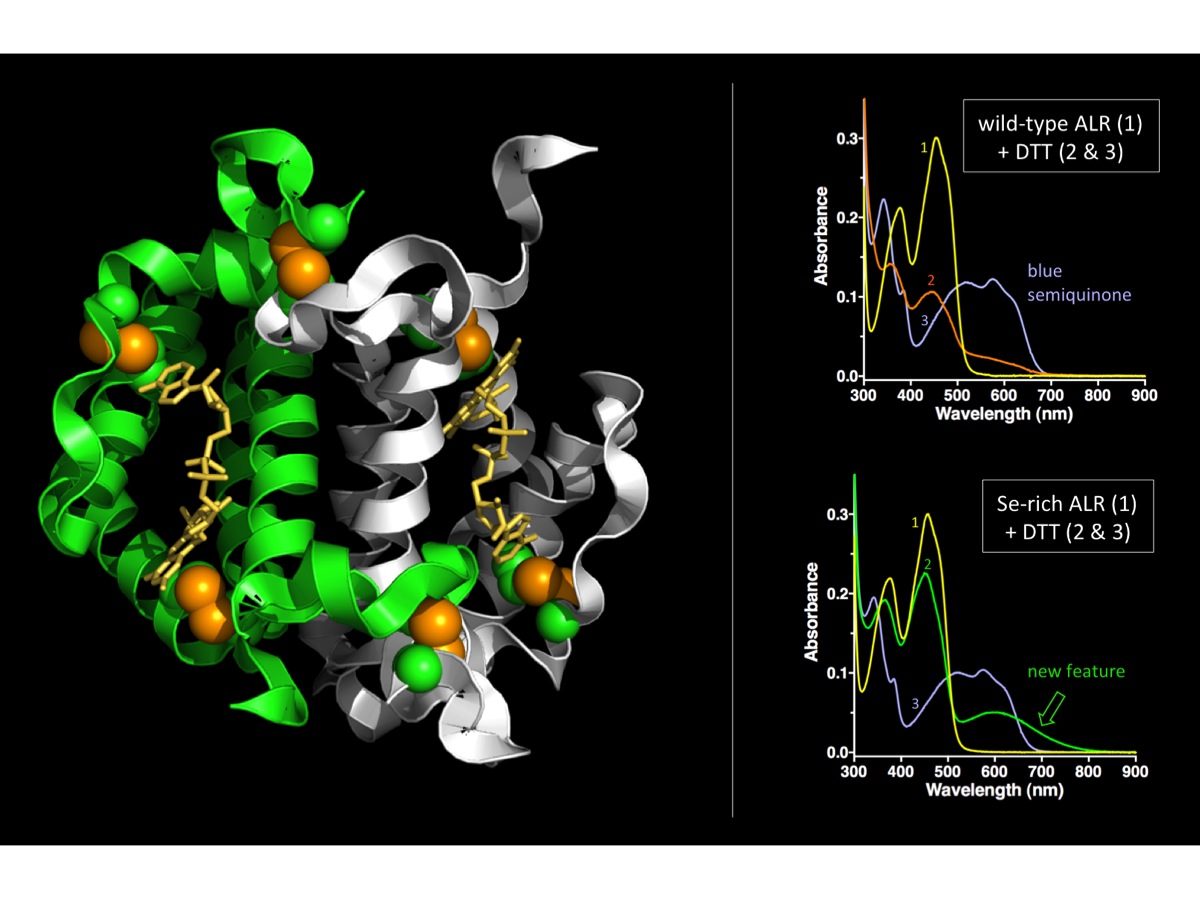 The right panels show the reduction of the enzyme by the model substrate dithiothreitol (DTT) monitored through the changes in the flavin spectrum. (Top panel is wild-type ALR; bottom panel is Se-rich ALR. Note the new long-wavelength absorbance feature that intervenes before the accumulation of the blue semiquinone).
The right panels show the reduction of the enzyme by the model substrate dithiothreitol (DTT) monitored through the changes in the flavin spectrum. (Top panel is wild-type ALR; bottom panel is Se-rich ALR. Note the new long-wavelength absorbance feature that intervenes before the accumulation of the blue semiquinone).
In sum, the work of Rozovsky and coworkers should help to put selenium into the biological NMR toolbox – as an environmental probe, and for the characterization of redox and ligation states. While attention has been drawn to the differences in reactivity between sulfur and selenium – they really do share lots in common.
So if you want to try an NMR-active surrogate for sulfur in your favorite protein … give 77Se a spin!
Notes
1) The paper: Stephanie A. Schaefer, Ming Dong, Renee P. Rubenstein, Wayne A. Wilkie, Brian J. Bahnson, Colin Thorpe and Sharon Rozovsky. 77Se Enrichment of Proteins Expands the Biological NMR Toolbox. Journal of Molecular Biology (2012 [PubMed].
2) Sharon Rozovsky’s website is here
3) Of course genetic incorporation of Se provides for site specificity – but to do this wholesale for a multicysteine-containing protein would generally require an unacceptable number of mutations to generate SECIS elements for each position.
4) In terms of “disulfides” 90% selenium incorporation corresponds to 1% -S-S-, 18% -S-Se- (or -Se-S-) and 81% -Se-Se-. So the majority of crosslinks in the selenium-ALR structure depicted in the Figure represent diselenides.
5) The PDB coordinates for the Selenium-rich form of ALR are to be found here
