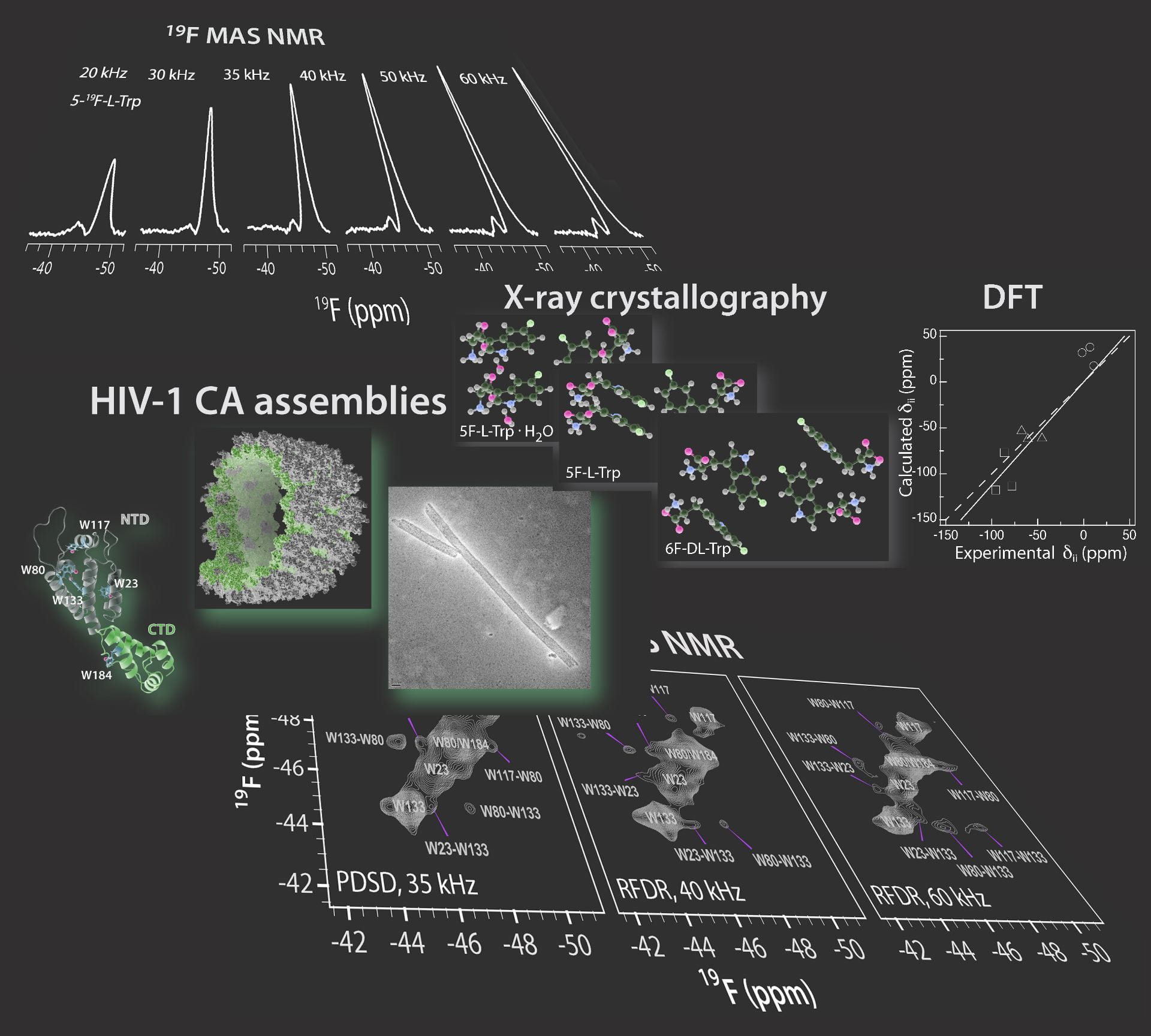
HIV-1 CA capsid assemblies 
Within the HIV-1 virion, the RNA genome is packaged inside a proteinaceous shell, the HIV-1 capsid core, comprised of 1000-1500 molecules of the capsid protein. HIV-1 capsid plays multiple roles in viral replication, and inhibition of capsid assembly is an attractive target for therapeutic intervention. The inherent structural plasticity of the conical HIV-1 capsid has hampered efforts to experimentally determine its structure. Our group uses cutting edge magic angle spinning (MAS) NMR spectroscopy to study HIV-1 capsid and its assemblies with cellular host factors and inhibitors with atomic resolution. We integrate MAS NMR with cryo-EM, MD simulations and QM/MM calculations to get atomic-resolution insights into structure and dynamics of capsid assemblies.
We recently determined the atomic-resolution structure of the HIV-1 capsid protein (CA) tubes, by MAS NMR and data-guided molecular dynamics simulations. The structure of the CA tubular assembly provides critical information, including conformations of the functionally important regions, which were inaccessible from the X-ray and cryo-EM structures reported previously, due to static conformational heterogeneity or motions. The structure provides important guidance for the design of novel capsid inhibitors. |
|
HIV-1 maturation
An essential step in the HIV-1 lifecycle, necessary for generating infective virions, occurs upon virus release from an infected host cell and involves processing of Pr55Gag (Gag) into its constituent domains (MA, CA, SP1, NC, SP2, and p6) by the viral protease. This sequential cleavage cascade is termed maturation. The final step in the maturation cleavage cascade is the removal of the SP1 peptide from CA, which triggers rearrangement of the immature lattice to form the final mature conical shape. The details of this conformational rearrangements remain elusive. Maturation inhibitors, such as Bevirimat, PF-46396, and their analogs, are thought to bind to the CA-SP1 junction, thereby blocking the SP1 cleavage and stabilizing the immature lattice. Our group integrates MAS NMR spectroscopy with other experimental and computational structural biology techniques to study the structure and dynamics of the maturation process and interactions with inhibitors. |
|
HIV-1 dynamics
Conformational plasticity and stability of the HIV-1 capsid are directly linked to its dynamic properties. The HIV-1 capsid is surprisingly dynamic, as we have discovered in our multiple studies. Segment- and residue-specific dynamics within individual CA chains appear to play diverse roles in the capsid. Millisecond-timescale motions of the flexible linker region fine-tune the relative NTD-CTD orientations. The CA flexibility conferred by the linker dynamics was evidenced in multiple cryo-EM structures and determines the variable curvature. Motions of residues in the CypA-binding loop that occur in the nano- to microsecond regime are connected to the dynamic regulation of CypA binding and the capsid’s escape from CypA dependence. Furthermore, the capsid’s interactions with cellular host factors CypA and TRIM5α are regulated by dynamics. TRIM5α binding to the capsid induces global rigidification and perturbs key intermolecular interfaces essential for higher-order capsid assembly. Dynamics are also essential in capsid maturation: quenching motions interfere with the maturation process. To study these processes, we integrate MAS NMR, MD simulations, and QM/MM calculations. |
|
Cytoskeleton and associated proteins
Microtubules, actin, and their associated proteins play essential roles in maintaining cell structure, organelle transport, mitosis, cell motility and cell division. Microtubules and actin filaments are highly regulated by hundreds of microtubule-associated proteins and actin-binding proteins, respectively. The complexes of cytoskeletal filaments and their various binding partners are important structural targets to elucidate cytoskeletal dynamics in cells. Structural characterization of these assemblies is challenging due to their inherent insolubility and lack of long-range order. We employ MAS NMR to study microtubule- and actin- protein assemblies. |
|
MAS NMR methods development
All of the ongoing projects in our group require development of new solid-state NMR methods. We have developed new experiments for i) studies of structure, dynamics, and intermolecular interfaces in protein assemblies at fast MAS frequencies (40-110 kHz) where conventional techniques do not work; ii) measurement of anisotropic NMR parameters for quantitative analysis of dynamics in proteins and protein assemblies; iii) 19F fast MAS NMR and 19F DNP-enhanced MAS NMR; iv) determination of relative CSA and dipolar tensor orientations by correlation spectroscopy at magic angle spinning; v) distance measurements between dipolar coupled spin-7/2 – spin-1/2 nuclei.
|
|